
The May (2008) issue of the Institute for Creation Research‘s monthly newsletter Acts & Facts contains an article by the current President of ICR, Dr. John Morris, titled “Evolution’s Biggest Hurdles“. The article is ostensibly about enumerating unsolved questions in evolutionary theory but instead what it does is highlight a deeply rooted set of character flaws in the “creation science” movement and its leaders: intellectual laziness (and/or dishonesty) combined with a lack of basic scientific literacy and colossal hubris.
While I understand that this article is a relatively short, non-technical piece, this does not in my opinion excuse the glaring omissions of relevant evidence about its supposed subject. Nor does the fact that Dr. Morris has a background in geological engineering forgive the zoological ignorance it displays. I am not a zoologist. I don’t have any degrees, but still I was able to immediately spot some of the rather glaring zoological errors and omissions in what Dr. Morris wrote. For someone who has been involved in the creation/evolution debate as long as Dr. Morris has it is difficult to fathom how he could not be better informed on such basic issues.
Morris: The theory of evolution has numerous problems, some of which are absolutely enormous and for which no adequate solution has even been proposed. The biggest problem comes right at the beginning with the supposedly spontaneous generation of life from non-life. Neo-Darwinian scientists admit this, recognizing that proposed evolutionary scenarios do not model reasonable conditions on earth, and could not have produced anything like the complex life we see all around us–even single-celled life.
I guess we’re going to keep repeating this until gets through their crania; the origin of life (abiogenesis), while it is related to the evolution of life, is nevertheless a separate question. Our current lack of a complete accounting of the process by which life originated has absolutely zero effect on evolutionary theory.
“Learn it. Know it. Live it.“
As for the complexity of living things and abiogenesis, I am unaware of any scientist working on the problem who believes that the first living thing was anywhere near as complex as a modern bacteria. Rather they think the first life forms would have been much simpler.
Also who are these supposed “Neo-Darwinian scientists” (who would not be likely to be working on abiogenesis) Morris is talking about? If he is going to make these sorts of claims it is incumbent upon him to cite specific examples. And which scenarios for “conditions on earth” are these mystery scientists admitting are not reasonable models?
This whole section is nothing but an unsupported a straw-man argument. But of course it isn’t intended as a serious scientific argument of any sort, it is really just a bit of fluff meant to nurture the prejudices of the faithful.
Morris: The second-biggest problem involves the development of complex invertebrates, animals without a backbone, from single-celled life. How did this transition occur? A robust fossil record of one-celled life has now been found, and of course a truly abundant record of marine invertebrates can be discovered everywhere, from clams to sponges to jellyfish to starfish, etc. The “explosion” of life in the Cambrian system of strata continues to baffle evolutionists, for there is no record showing a transition from tiny single-celled life to complex invertebrates. There are innumerable fossils of invertebrate ocean bottom life, even those with no hard outer shell, but no ancestors of these invertebrates have been identified.
While it is true that there are still plenty of unanswered questions remaining about how multicellular eukaryotic organisms (metazoans) evolved from single celled eukaryotic organisms and that fossil evidence is generally lacking in this area, we are not, as Dr. Morris would have his readers believe, completely in the dark.
If we don’t have a lot of fossils we can look to living organisms to see if any of these might fill in some of the missing pieces of the puzzle using comparative anatomy, embryology and molecular biology.
The origin of metazoans: Living clues
There are a number of different types of single celled organisms that come together to live in cooperative colonies that blur the distinction between single celled and multicellular life.
For example there is the unicellular slime mold Dictyostelium, the so-called “social amoebae” which ordinarily lives as a single celled organism but under certain conditions (a shortage in food) they group together into a slug-like form and act almost as a single multi-celled organism.
From the plant world there is Volvox a type of colonial green algae which grow into beautiful spheres (some large enough to see with the naked eye) consisting of many hundreds or even thousands of individuals held together by strands of cytoplasm.
choanoflagellates
However the evidence from morphological and molecular studies both point towards a group called choanoflagellates as being the group of single celled organisms most closely related to metazoans (Brooke & Holland 2003) (King 2004). Some choanoflagellates live entirely as single celled organisms while others form colonies of various configurations (flat, spherical, in blobs of gelatinous secretions) (Nielsen 2008). Below is an illustration of a stalked form of colony.
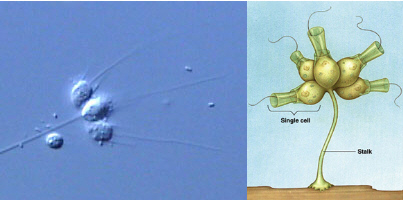
See original photo here and diagram here.
Porifera
Next we have the sponges, whose interiors are lined with cells called choanocytes which are dead ringers for, you guessed it, choanoflagellates.
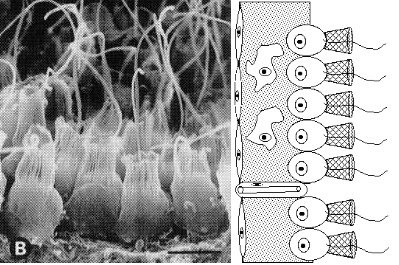
On the left a scanning electron microscope image of choanocytes from a demosponge (Nielsen 2001, p.15) and on the right a diagram of sponge choanocytes in relation to other cell types.
Sponges, having no specialized organs and only a dozen or so cell types, are one of the most morphologically simple types of metazoans. They are also one of the earliest to appear in the fossil record (late pre-Cambrian). Molecular studies also indicate that sponges are basal within the metazoans.
The study of sponge systematics is from what I can gather still in a state of flux, however there seems to be an emerging consensus that they are not monophyletic but rather should be divided into at least three separate clades: Hexactinellida, the Demospongia, and the Calcarea, perhaps more.
Some molecular studies (Borchiellini et al 2001) seem to indicate that the Calcarea are the most closely related to other metazoans. On the other hand some molecular and morphological evidence suggests that a group of sponges called Homoscleromorpha might be more closely related to other metazoans than calcarean sponges. Perhaps the most telling morphological character present in Homoscleromorpha is the epithelium, something lacking in other sponges but found in all other metazoans.
Homoscleromorpha are usually classified as belonging to the demosponges, but I have also seen them placed in their own separate clade. So stay tuned on sponges.
Placozoa
Another organism of interest is the Phylum Placozoa, known from a single living genus Trichoplax. It was originally discovered not in the wild but living in an aquarium in the 1880’s. Their bodies being basically flattened amorphous blobs made up of only four basic types of cells making them the simplest known multicellular organisms. They move and behave much like giant multi-celled amoebas. They can reproduce by fission and apparently through sexual reproduction as well, though little is known about the details of the process. In fact much about them remains a mystery.
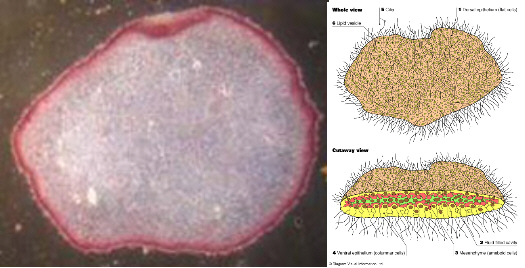
See original photo here and diagram here.
Trichoplax has a sealed epithelium and unlike sponges has extracellular digestion. It feeds by ‘climbing’ on top of its prey (algae etc.) and then digesting it with the cells on their undersides.
Their relationship to other basal metazoans remains uncertain, but what is fairly certain is that these simple creatures probably have a lot to tell us about early metazoan evolution.
Cnidarians
Yet another group at the base of the metazoan family tree are the Cnidarians (jellyfish, sea anemones, corals) which are, along with sponges, among the earliest multicellular organisms found in the fossil record (also late pre-Cambrian). Cnidarians radially symmetrical and develop from two primary germ layers (diploblasty). They basically have a cup or bowl shaped anatomy with a single opening for use as both mouth and anus, and the inside of the ‘cup’ forming the gut where food is digested. As with placozoans digestion is extracellular.
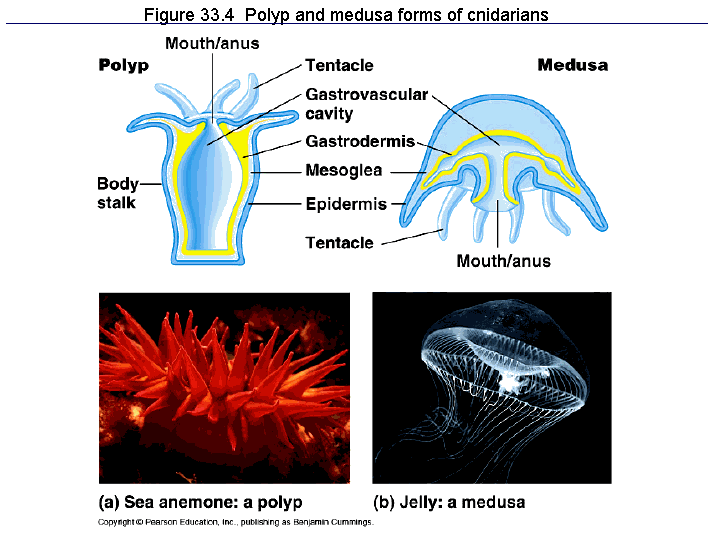
This diagram can be found on this web page.
Some recent studies suggest that some cnidarians exhibit the morphological rudiments of bilateral symmetry and having similar Hox genes to bilaterally symmetrical metazoans.
Acoelomorpha
Finally we have the most basal living bilaterally symmetrical animal the acoelomorphs that closely resemble the Playtyhelminthes (flatworms) which until fairly recently they were classified as (Ruiz-Trillo et al 1999).
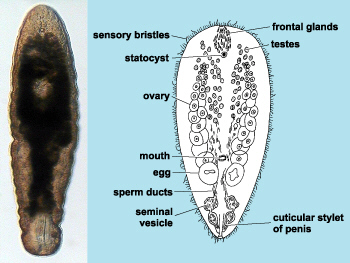
See original photo here and diagram here.
While they are bilaterally symmetrical, their nervous system is a decentralized network similar to that of radially symmetrical cnidarians (Valentine 2004, p. 240). They have only one orifice for both eating and expelling waste just as cnidarians do and they also resemble somewhat the planula larva of some cnidarians.
There is more information available on these and other organisms that fill in the gap between single celled organisms and more complex bilaterally symmetrical metazoans like arthropods, mollusks, and vertebrates, much of it could be learned by picking up a standard zoology textbook, or even by doing a few Google searches.
So the question is this: is Dr. Morris ignorant about this information and too lazy to try to learn about it or does he know about it but simply chose not to inform his readers about any of it and thereby lie by omission?
Acoelomorph update (3-10-2011): Recently there was a study published in the journal Nature (Philippe et al. 2011) which, based on molecular evidence, suggests that rather than being basal within bilateral metazoans, acoelomorphs may instead be basal deuterostomes that have secondarily lost their anuses and reverted to a single orifice. However, as of this updating the claim remains controversial, so stay tuned on acoelomorphs.
For more details on the recent acoelomorph study and the controversy see: “A Can of Worms” by Amy Maxmen.
The Fossil Record and the origin of metazoans
Dr. Morris brings up the so called Cambrian “explosion” (as creationists love to do) which he says “baffles” evolutionists. However if evolutionists are curious about the Cambrian radiation it is because, unlike creationists, scientists take the evidence seriously and wish to understand what actually happened during this part of the history of life on earth.
Paleontologists have worked for over a hundred years collecting Cambrian (and pre-Cambrian) fossils, publishing countless papers detailing what they have found and trying to make sense of it in light of what we know about evolutionary processes and the history of life since that time. Despite all this there is still not a consensuses on exactly what occurred and debates on the timing, mechanisms involved, and whether there really even was an ‘explosion’ at all continue with no quick resolution in sight (Fortey et al 1996) (Cartwright & Collins 2007).
The quest to understand what happened is often made problematic by the fact that the fossil record is far from complete and fossils, when found, are often difficult to interpret due to poor and limited preservation. Another important source of data is molecular evidence, but just as with fossils our current knowledge is limited and so we often get equivocal results. So the search for more evidence and the debates over that evidence continues slowly putting the pieces of the puzzle together.
To give you an idea of how incomplete the fossil record can be in this case it should be noted that three of the five animal groups I mentioned in the previous section don’t have any fossil record at all. Choanoflagellates (Porter 2006) (Lipps 2006), Placozoans (Valentine 2004, p. 233), and Acoelomorphs (Valentine 2004, p. 240) have never been found as fossils. Clearly the ancestors of the living species of these groups must have existed since at least the late pre-Cambrian but being small and soft bodied they left no trace (discovered so far) of the millions upon millions of generations that have lived and died since that time.
Most or all of the organisms that would be ancestral to those found in the Cambrian radiation were likely very small, soft-bodied, creatures rarely preserved as fossils (and even when they are it is often difficult to make sense of them) and despite what Morris implies, fossils preserving soft-bodied organisms are hardly “abundant”. Instead they are relatively rare paleontological treasures largely found in a few special locations like the Burgess shale of Canada and the Maotianshan shales of China. .
Creationists on the other hand are only interested in how they might be able to fashion the Cambrian radiation into a rhetorical cudgel with which they can attack evolution. They have no explanation for the Cambrian radiation, or any other part of fossil record, except for their ridiculous Flood geology which falls apart after an even cursory examination of the facts.
For example, the fossil record we have for marine invertebrates (from the Cambrian to the Pleistocene) is a time ordered pattern of change which cannot be explained by the Flood geology (espoused by both Morris and his organization) and this was recognized by scientists long before evolution was widely accepted in the scientific community (see Ritland 1981 & 1982 in suggested reading below).
The following is a table from Morton & House (2001) listing the order of appearance of various fossils from the pre-Cambrian to the end of the Cambrian period:
Table 1. Cambrian Chronology (Except where otherwise noted, data from ref. 55)
Million Years Ago (myr) Event
495 End of Cambrian
511 Wheeler Formation Fauna
518 Burgess Shale Formation
520 First Hemichordates
523 Kaili deposits
526 First Wiwaxia found in Mount Cap Formation
527 First trilobite
530 Chengjiang fauna–stem groups deuterostomes, first Ctenophores
531 First Cephalochordates, first chordates
533 Sirius Passet fauna priapulids, first annelids
534 First brachiopods, first molluscs
535 First arthropods
536 First Lobopodians/Anomalocarians
537 First nematodes, Halkieriids and echinoderms
538 Abundant Shelly fossils
541 Skeletonized foraminifera
543 Small shelly fauna
545 Cambrian/Precambrian Boundary- Cambrian above/Precambrian below
546 Cloudina stem group ecdysozoans
548 First stem group lophotrochozoans
550 Advanced Ediacaran fauna–Spriggina [trilobite precursor]
553 First anthozoan
555 Kimberella (first mollusc forerunner)
560 Earliest fungi, earliest stem-group cnidarian
563 Mistaken Point faunas
568 End of ice age?
571 First sponge (Doushantuo)?
591-610 Varanger-Marinoan glaciation
610 Oldest Ediacaran fossils Mackenzie Mountains, Canada
680 Oldest accepted worm tracks
1700 Oldest multicellular fossils
[Table from Morton & House 2001]
The fossil record indicates that the first few billion years of life on Earth was dominated by prokaryotic single celled organisms (bacteria) like the blue-green algae that create stromatolites. The oldest single celled eukaryote fossils occur about 2 billion years ago (Waggoner 2001). The first taxonomically resolvable multicellular eukaryote fossils (the red algae Bangiomorpha) do not occur until 1.2 billion years ago (Butterfield 2000) and it is not until the last 600 million years or so that more complex multicellular animals evolved.
Morris specifically mentions four types of organisms; clams, sponges, jellyfish and starfish implying that these all appeared during the Cambrian radiation. This is not the case.
Both Sponges and jellyfish are known from pre-Cambrian strata and thus predate the radiation. “Clams” or rather bivalve mollusks are indeed known from the Cambrian but only from extinct types (so half a point for Morris there I guess) but ‘Starfishes’ (Class Asteroidea) fossils do not appear in the fossil record until the Ordovician.
The same often holds with regards to the host of organisms he didn’t mention. For example while there are many types of arthropod fossils found in Cambrian strata there are none of the familiar decapod crustaceans (crabs, lobsters, and shrimps) or hexapod insects found in the fossil record until the Devonian period, about 100 million years after the Cambrian radiation.
Intermediate forms in the Cambrian
While the fossil record is far from complete, there are fossils that add more pieces to the puzzle. Many late pre-Cambrian and Cambrian fossils are difficult to pigeon-hole into extant animal groups. Some appear to have no living representatives (Nectocaris, Odontogriphus etc.) while others appear to be either basal members of existing taxa (Sanctacris, Haikouella etc. ) or intermediate between them and as with the pattern of the fossil record creationism has no logical explanation for the existence of organisms that are difficult to classify.
A few well know examples of possible intermediates (NOTE: no claim of direct ancestor descendant relationships between the genera or phyla listed below is intended here):
- The Dinocaridida (Dinocarida) which appear to have both onychophoran and arthropod characteristics. Examples are Kerygmachela, Opabinia, Parapeytoia and Anomalocaris
- Halkieria which appears to have characteristics of brachiopods, annelids and mollusks.
An evolutionary relationship between onychophorans and arthropods is also supported by molecular evidence, as is that between brachiopods, annelids and mollusks (Dunn et al 2008).
Of course I don’t expect that any of this information would convince a creationist but Dr. Morris’s characterization of our state of knowledge about the transition from single celled organisms to complex metazoans (even if he personally doesn’t agree with it) is not even remotely fair or accurate. An intellectually honest critique of the mainstream science position would have to point this sort of information out if only to explain why it supposedly fails to make the case.
However Morris’s comments in his article don’t even rise to the level of being a mischaracterization of what mainstream science has to say on the origin of metazoans. He doesn’t give his readers the slightest hint that anything like this evidence is out there (again even if he disagrees with it or its interpretation), instead, out of ignorance, laziness or malice, he says nothing about it.
Morris: A third huge problem lies in the next step required by evolution. Fish, thought to be the first vertebrates, must have evolved from invertebrates, but again there is no record of this transition. “How this earliest chordate stock |i.e., early vertebrates| evolved, what stages of development it went through to eventually give rise to truly fishlike creatures, we do not know.”
Despite what Morris implies by in his bracketed interjection in this quote (“|i.e. early vertebrates|”), not all chordates are vertebrates. Vertebrates are a sub-phylum of chordata, so all vertebrates are chordates but not all chordates are vertebrates.
Molecular studies indicate that invertebrate chordates are more basal than vertebrates, thus the common ancestor of all chordates was probably more similar to the invertebrate chordates than to vertebrates. Furthermore the earliest chordates in the fossil record are invertebrate chordates (more on these in a moment) and possibly jaw-less ‘fish’ something like a hagfish or lamprey.
Most fish you’ve heard of, eaten or kept in a fish tank (goldfish, trout, halibut, clownfish, guppies, barracuda, bass etc.) are ray finned fish i.e. teleosts. The first teleost fish, which dominate both fresh and salt water environments today, don’t appear in the fossil record until the Late Triassic period (some 220 mya).
Before going on I want to address the fact that Dr. Morris gives one, and only one reference for this entire article The Fishes (1964) by Francis Downes Ommanney. What is stunning about this, is that of the huge mass of peer reviewed scientific literature and textbooks that have been produced on the subject, Dr. Morris chose a 44 year old popular level book from the Time-Life Nature Library series.

This book is not part of the scientific literature and this is not simply my (non-professional) opinion. Here is a quote from a contemporary review of the book published in the journal Science by the late ichthyologist Giles W. Mead (1928-2003) then of the Museum of Comparative Zoology at Harvard University:
Those who wish a superficial account of the biology of fishes may find this book enjoyable. … The errors are few, and these are chiefly the result of oversimplification without qualification. … But the complete absence of documentation makes verification impossible, an omission which also makes the volume a dead end rather than a means of access to the literature of ichthyology and related subjects. The one-page bibliography is inadequate. Even access to source material through acknowledgment is lacking. …It will be informative to the lay reader and perhaps useful to the high school biology student. But since it lacks both depth and documentation, it can serve no higher scientific or educational purpose. (Mead 1963, emphasis mine)
Dr. Morris chose this as his single reference but I don’t even know for sure that he looked it up independently while doing “research” to write this article. You see the quote he gives from The Fishes appears on page 180 in the book That Their Words May Be Used Against Them (1997) written by Dr. Henry M. Morris (1918-2006), John Morris’s father. This book, subtitled: Quotes from Evolutionists Useful for Creationists, is a compilation of disembodied quotes from mainstream scientists, quacks and creationists.
So Dr. [John] Morris may have simply opened up the out-of-context-quotation database and stuck one he thought might be appropriate into his piece without even bothering to pull The Fishes off the shelf.
I also note a typo in the reference. The quote from Ommanney appears on page 60 of The Fishes, not page 64 (maybe ICR could fix that in the online version). [I know, I know, I’m just piling on now…]
Morris: Over the years nearly every invertebrate has been proposed as the ancestor, but each suggestion has only been in vogue for a time. … One proposition receiving attention these days is that echinoderms were the creature that evolved into fish.
Chordates (never mind “fish”) share a close common ancestor with echinoderms; they did not evolve from them, at least not any of the existing types. Existing echinoderms, with their secondarily developed radial symmetry, are too highly derived to be the ancestors of chordates. It has been suggested by some scientists (Jefferies 1997) that certain extinct forms of basal echinoderms (the Stylophora for example) might have given rise to chordates but this hypothesis remains highly controversial (Donoghue et al 2003, p. 194).

Image source Palaeos.com
Generally modern biologists and paleontologists shy away from labeling groups as being ancestral to one another (especially living groups) and instead refer to ‘sister groups’ or ‘sister taxa’ when talking about closely related groups. It this case ambulacraria (echinoderms + hemichordates) and chordates could be sister groups.
It is technically true that various invertebrate groups other than echinoderms have been proposed as being closely related to chordates, but these other scenarios are primarily from the 19th century. However the current consensus on there being a close relationship between echinoderms and chordates is not merely a recent fad, as Morris seems to imply.
It was recognized in the late-19th century that the larva of certain echinoderms and the hemichordates (acorn worms) were strikingly similar and given that hemichordates were at that time classified as invertebrate chordates this was seen as a strong evidence linking echinoderms and chordates.
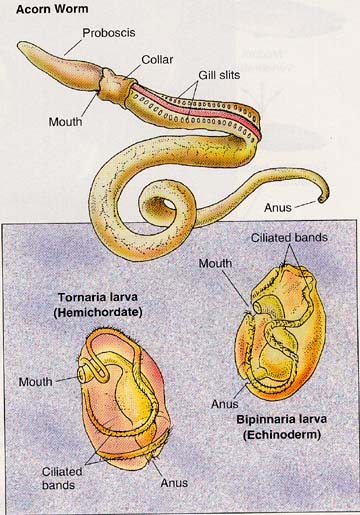
Above is an illustration comparing the larva of hemichordates to a bipinnaria echinoderm larva. It can be found on the Bethel University (St. Paul, MN) web site.
Here are a few quotes from zoology books from over the last 120 years which discusses the evidence linking echinoderms and chordates via hemichordates (note: the genus Balanoglossus in the following quotes refers one of the hemichordates just discussed):
I prefer to look upon the Proto-Chordata as having arisen from a more generalised form than either the Nemerteans or the Annelides. Their point of origin was certainly very far back, and may have been even lower down on the vermean axis than its position in the table. From the striking resemblance between the respiratory organs of Balanoglossus and those of some of the lower Chordata it is extremely probable that the Proto-Chordata started from ancestral Enteropneusta, and it has been already seen (page 40) that the Enteropneusta and the Echinodermata probably possessed a common ancestor outside the vermean series, consequently the Proto-Chordata, the Enteropneusta, and the Echinodermata most probably diverged from the main axis together as a branch which very soon gave off the first ancestors of the Chordata in one direction and the primitive Echinoderms in another. (Herdman 1885, p. 57)
The evidence in favor of an alliance between Vertebrates and Echinoderms (sea urchins and starfishes), through the intervention of Balanoglossus, seems, however, to be steadily gaining ground. Mr. Willey, for instance, remarks that while it is probable that the proximate ancestor of the Vertebrates was a free swimming creature, intermediate in structure between an ascidian larva and the lancelet, the ultimate or primordial ancestor may be assumed to have been a worm-like animal, with an organization approximately on a level with that of the bilaterally symmetrical progenitors of the Echinoderms. (Lydekker 1901, p. 2953)
We have now passed in review all the points of resemblance between the Echinodermata and the Chordata and we have seen that they all, except possibly the last, are concerned with fundamental, not superficial, traits. Some of them link the Echinoderms to all the Chordates or to all except the Tunicata, e.g. the relations of the central nervous system and the general relations of the coelomic sacs; another they hold in common only with Vertebrata, viz. the presence of calcified plates in the mesoderm of the body wall; a third – the sinistral position of the mouth – is found again only in Amphioxus; and lastly there is the striking resemblance to Enteropneusta [hemichordates–T.B.] by the tornaria larva. The accumulated weight of these facts is overwhelming and leaves us no choice but to consider not only that the Chordata are the nearest allies of the Echinodermata, but that the Echinodermata are of all Coelomata the nearest to the Chordata. (Sedgwick 1909, p.165)
Echinoderm Theory. – In spite of their aberrant organization, echinoderms afford the best clue to chordate ancestry. The resemblance between the bipinnaria larva of echinoderms and the tornaria larva of hemichordates, as well as the striking similarity in the mesoderm formation in the two groups, indicates some relationship somewhere phylogenetically. Although adult echinoderms have radial symmetry, their larval forms are bilaterally symmetrical.
In some other respects there are patterns of similarity between the echinoderms and the chordates. Both have indeterminate cleavage; that is, each of the early blastomere cells have equivalent potentialities with each of the other blastomeres. The coelom in each is formed by an outpushing of the archenteron. Also, the anus arises from the blastopore and the mouth has a secondary origin. At one time biologists gave some attention to a possible affinity between the chordates and the composite group, annelids-arthropods-mollusks. But among the members of these phyla there is determinate cleavage and a different mode of mesoderm formation, the coelom is hollowed out of mesoderm bands, the blastopore forms the mouth, and the anus arises secondarily. (Hickman 1955, p. 377, Emphasis in original)
While hemichordates have since been re-classified into their own separate phylum (as a sister group to echinoderms) the molecular evidence discovered since the 19th century has only strengthened the case for the common ancestry of echinoderms, hemichordates and chordates. Bringing up the fact that other groups besides echinoderms were at one time considered as close relatives to chordates is clearly a red herring meant to imply that the basic phylogenetic relationship of echinoderms and chordates is still an open question. It isn’t.
Morris: As Dr. Duane Gish–former Senior Vice President at ICR and a well-known creation scientist–likes to say, if evolution can’t derive either invertebrates from single-celled life, or vertebrate fish from invertebrates, it is “dead in the water.”
This brings us back to a discussion of the invertebrate chordates which Dr. Gish and Dr. Morris act as if they have never heard of. Actually I’m sure they have, they just choose to pretend that they don’t exist.
There are two living groups of invertebrate chordates, the cephalochordates (commonly called the ‘lancelet’ and in older textbooks ‘amphioxus’) and the urochordates (sea squirts, salps and larvacea).
Lancelets are fairly fish-like but they lack any paired fins, jaws, complex eyes and even a proper brain. They live with their ‘heads’ sticking out of the sea floor, filtering food particle out of the water with their gills.

A diagrammatic illustration of a cephalochordate. Image from the online version of Harold L. Levin’s: The Earth Through Time (8th Ed.).
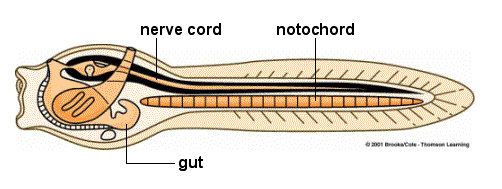
A diagrammatic illustration of a urochordate larva. The original image can be found on the Teaching Resources Center of the UC Davis web site.
The free swimming sea squirt larvae eventually settle down, ‘nose’ first, and metamorphose into the much different sessile adult form. Like the lancelet, adult sea squirts make their living by filtering water for food. It was traditionally thought, based on comparative anatomy that cephalochordates were more closely related to vertebrates and that urochordates, with their significant morphological differences, represented more basal chordate characteristics. But recently there have been some molecular studies which indicate urochordates may be more closely related to vertebrates than cephalochordates. If this turns out to be the case then it would indicate that the cephalochordate/vertebrate body plan is basal and that urochordates are fairly derived, having evolved on their own unique direction.

The larva of a lamprey (a basal vertebrate) for comparison to cephalochordates and urochordates larvae.
Early Fossil chordates
The earliest chordates found in the fossil record are from the Cambrian and generally resemble cephalochordates in structure. Here are a few examples:


Pikaia gracilens, from the Burgess Shale of Canada. The photo on top is from the Smithsonian Institute web site and the drawing below is from this (French language) site.

Haikouella lanceolata from the Yunnan Province of China. Both the photo and diagram are from the Palaeos web site.
Below is a possible basal vertebrate (similar to a hagfish or lamprey) Myllokunmingia fengjiaoa also from the Yunnan Province of China.

Original picture found in online version of Richard Dawkins’ book The Ancestor’s Tale (2004).
There is also an enigmatic fossil group called vetulicolians (also from the Cambrian strata of China) which have been interpreted by some workers as being basal deuterostomes and perhaps ancestral to both chordates and echinoderms.

“The deuterostomes, and possible places for vetulicolians in the evolutionary scheme of events” from (Gee 2001). Click on image for larger version.
These early chordates are joined in the fossil record during the Ordovician by the (jawless) pteraspidomorphi, the armored and still jawless osteostraci during the Silurian. By the end of the Silurian jawed fish finally make an appearance in such forms as the acanthodians (the so-called ‘spiny sharks’).
The first chondrichthyes (the group to which sharks and rays belong) and actinopterygii (ray-finned fish) appear in the Devonian (though there is evidence of a possible Silurian actinopterygian). As noted earlier the dominate group of fish today, the ray-finned teleosts do not make showing in the fossil record until the Triassic.
Once again Morris gives his readers no hint of any of this evidence (even if he disagrees with it or its interpretation), and once again one is left to wonder whether it is out of ignorance or malice.
Morris: Now, echinoderms usually don’t look anything like fish. Their ranks include sea squirts, tunicates, and starfish. These do have a pseudo-spine with a central supportive notochord and a tubular nerve chord, features that are somewhat present in vertebrates, especially in the embryonic stage.
There are just a few teeny tiny little nits to pick here, as you may have guessed given what we just talked about above.
First, the ranks of echinoderms (phylum Echinodermata) cannot include both sea squirts and tunicates because sea squirts are tunicates. They are the same animal. Sea squirts is a common name for tunicates.
Second, the much more important reason tunicates cannot be counted amid the ranks of echinoderms is because they simply are not echinoderms. They are chordates (sub-phylum urochordata), one of two living types of invertebrate chordates I’ve been going on about. Morris’s mistake here is akin to saying that the ranks of insects include the squid and octopus.
Third, despite what Dr. Morris says, echinoderms, like starfish or sea urchins, do not at any point in their lives have a notochord or a dorsal hollow nerve chord, these are chordate characteristics. Continuing our insect/squid analogy, this is the equivalent of saying that that insects have tentacles and radula (as opposed to jointed legs and mandibles). Furthermore, these characteristics are not merely “somewhat present” in chordates (including vertebrates) they are two of the defining characteristics of the phylum.
This is very basic stuff and frankly anyone who is this ignorant of such basic taxonomic distinctions is in no position to be criticizing evolutionary theory.
And yet Dr. Morris heads an organization whose very reason for being is to criticize evolution. What astonishing hubris it must take to make a career traveling about the world confidently declaring that practically the entire scientific community for the last 150 years is in gross error about the subject of evolution when, based on this article, Morris would apparently struggle to pass a basic zoology exam.
Morris: It seems that Ernst Haeckel’s theory of embryonic recapitulation is more extensive than once thought.
This has become a knee-jerk creationist syllogism whenever evidence for evolution from comparative embryology comes up. It usually goes something like this:
The evidence for evolution from comparative embryology = Haeckel’s drawings and/or Haeckelian recapitulation.
Haeckel’s drawings are, very inaccurate or deliberate frauds and the recapitulation theory has long been discredited.
Therefore the evidence from comparative embryology = deliberate fraud and long discredited theory.
This is all nonsense of course. Just because comparative embryology is involved doesn’t mean that Haeckel or his theories has anything to do with it.
Morris: It has been discovered that during embryonic development, certain features in the gut of the starfish bear similarity to a feature in vertebrate embryos. Further maturity yields a form and function far different from the vertebrate condition, yet this early formative pathway seemingly provides the hoped-for clue evolutionists need.
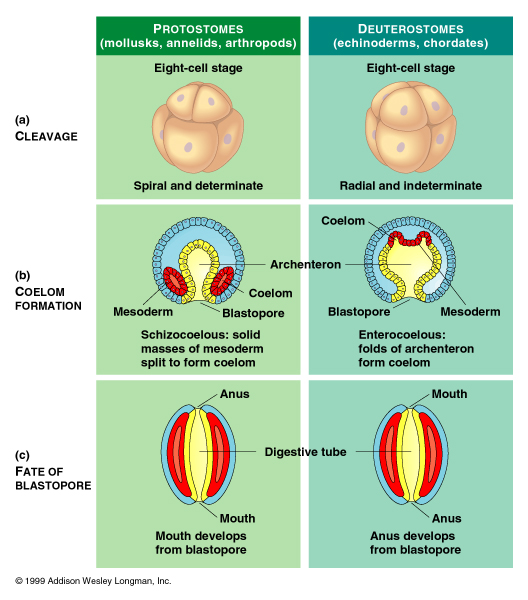
I can only guess at what Dr. Morris is talking about here as he writes in vague general terms and gives no references. What I assume he is referring to is the fact that both echinoderms and chordates are deuterostomes. That is, during embryological development of both these phyla the anus forms from a structure called the blastopore.
Click on image for larger, clearer, version, or here for source page.
While “further maturity” certainly does change the “form” of deuterostomes (quite radically in echinoderms) it does not change the basic “function”. Adult echinoderms and chordates both have anuses originally formed from the embryonic blastopore.
In both the Chordata and Echinodermata we find strong embryological evidence for the assumption that the blastopore became anus only and that the present mouth is a new pore. In every case described the anus is always either derived directly from the blastopore or else it opens at the spot where the blastopore has closed. Thus the Chordata and Echinodermata agree in their main axial relation to the plane of the blastopore, for, in each case it is at right angles to this plane. They also agree in various other respects. (Assheton 1916 pp.58-59)
Like the similarities between echinoderm and hemichordate larva this is not a recent discovery (as is evident from the above quote) and it was, and is, considered another strong bit of evidence for a close evolutionary relationship between echinoderms and chordates.
Morris: Is this proof that a starfish evolved into a fish that evolved into a human? It seems to me that they are grasping at straws.
No one that I am aware of is suggesting that “starfish” are the ancestors of fish or humans and in this case it would seem that it is Dr. Morris who is grasping at straws, apparently for use in creating straw-men of evolutionary theory to attack.
Morris: Without the assumption of evolution, without the neo-Darwinian scientist’s “certain knowledge” that vertebrates evolved from invertebrates, this flimsy link would not merit such attention.
This “flimsy link” is supported by evidence from comparative anatomy, embryology, paleontology and genetics. Dr. Morris’s support for his position is nothing more than his personal Biblical interpretation.
Morris: How much more scientific it is to recognize the God-designed diversity in creation, and not attribute all living things to a hypothetical common ancestor. Scripture teaches, “All flesh is not the same flesh: but there is one kind of flesh of men, another flesh of beasts, another of fishes, and another of birds” (1 Corinthians 15:39).
Simply repeating ‘God-did-it’ is scientifically worthless, Dr. Morris, as I am sure you’ve been told innumerable times. All you are doing is relabeling your ignorance and calling it an explanation and despite what you might like to believe scripture is not empirical evidence.
Conclusion
This article is a microcosm of the creationist intellect. False, misleading or incomplete, information used to disparage evolutionary theory presented with absolute confidence and authority. Exactly the sort of business that set me on a lifelong crusade against creationism.
How much of this is due to dishonesty (intellectual or otherwise) and how much is ignorance is often hard to tell. However, when one considers that A) Dr. Morris’s only job, as a leader in the creationist movement, is thinking, writing and talking about the subject of evolution, B) how much information is readily available at libraries and on the internet and C) that while observing and/or participating in countless debates during his career he must have heard many informed individuals talking about at least some of this information, it is difficult to believe that the problem is simply honest ignorance.
It is one thing for an individual to privately believe whatever they want; believing in YEC or disbelieving evolution. It is another thing entirely to stand up and confidently tell others that evolution is false, to imply that scientists are deceitful tools of evil and to claim that science really supports your YEC position, all the while using false, misleading, or incomplete information to dupe your audience.
Some suggested reading
Michael J. Benton (2004) “Vertebrate Origin“, Chapter 1 of Vertebrate Palaeontology (Third Edition).
Simon Conway-Morris (2003) “The Cambrian explosion of metazoans and molecular biology – would Darwin be satisfied“, International Journal of Developmental Biology 47: 505-515
Andrew H. Knoll and Sean B. Carroll – Early Animal Evolution: Emerging Views from Comparative Biology and Geology
Bruce S. Lieberman (2003) “Taking the Pulse of the Cambrian Radiation“, Integrative and Comparative Biology 43(1):229-237
Kieth Miller – The Precambrian to Cambrian Fossil Record and Transitional Forms
Glenn R. Morton and Ramsden House (2001) “Transitional Forms and the Evolution of Phyla“, Perspectives on Science and Christian Faith 53(1):42-51
Richard Ritland (1981) “Historical Development of the Current Understanding of the Geologic Column: Part I“, Origins 8(2):59-76
Richard Ritland (1982) “Historical Development of the Current Understanding of the Geologic Column: Part II“, Origins 9(1):28-50
J. William Schopf (2000) “Solution to Darwin’s dilemma: Discovery of the missing Precambrian record of life“, Proceedings of the National Academy of Science USA 97(13): 6947-6953
References
Assheton, Richard (1916) Growth in Length: Embryological Essays, Cambridge at the University Press
Borchiellini, C. et al (2001) “Sponge paraphyly and the origin of Metazoa”, Journal of Evolutionary Biology 14(1):171–179
Brooke, Nina M. & Holland, Peter W. H. (2003) “The evolution of multicellularity and early animal genomes” Current Opinion in Genetics & Development, 13:599-603
Budd, Graham E. (2003) “The Cambrian Fossil Record and the Origin of the Phyla“, Integrative and Comparative Biology, 43:157-165
Butterfield, Nicholas J. (2000) “Bangiomorpha pubescens n. gen., n. sp.: implications for the evolution of sex, multicellularity, and the Mesoproterozoic/Neoproterozoic radiation of eukaryotes”, Paleobiology 26(3):386-404 (abstract)
Cartwright, Paulyn & Collins, Allen (2007) “Fossils and phylogenetics: integrating multiple lines of evidence to investigate the origin of early major metazoan lineages”, Integrative and Comparative Biologyabstract) 47(5):744-751
Donoghue, Philip C. J. et al (2003) “The origin and early evolution of chordates: molecular clocks and the fossil record”, in Donoghue, Philip C. J. & Smith, M. Paul (Editors) (2003) Telling the Evolutionary Time: Molecular Clocks and the Fossil Record (Systematics Association Special Volume), Taylor & Francis Inc.
Dunn, Casey W. et al (2008) “Broad phylogenomic sampling improves resolution of the animal tree of life”, Nature 452:745-750 (abstract)
Fortey, R. A. et al (1996) “The Cambrian evolutionary ‘explosion’: decoupling cladogenesis from morphological disparity”, Biological Journal of the Linnean Society 57:13-33 (abstract)
Gee, Henry (2001) “On being vetulicolian”, Nature 414:407-409 (abstract)
Herdman, William A. (1885) A Phylogenetic Classification of Animals (for the Use of Students), Macmillan And Co. London
Hickman, Cleveland P. (1955) Integrated Principles of Zoology, The C. V. Mosby Company, St. Louis, MO
Jefferies, Richard (1997) “A defence of the calcichordates”, Lethaia 30:1-10
King, Nichole (2004) “The Unicellular Ancestry of Animal Development” Developmental Cell, 7:313–325
Lipps, Jere H. (2006) “Major Features of Protistan Evolution: Controversies, Problems and a Few Answers“, Anuário do Instituto de Geociências, 29(1):55-80
Lydekker, Richard (Editor) (1901) Library of Natural History, Vol. V, Sec. II
Mead, Giles W. (1963) “Life Nature Series: Two Subjects, Two Viewpoints: The Fishes”, Science (New Series) 140(3567):626
Morris, Henry M. (1997) That Their Words May Be Used Against Them, Master Books Inc. p. 180
Morris, John (2008) “Evolution’s Biggest Hurdles“, Acts & Facts 37(5):13
Morton, Glenn R. & House, Ramsden (2001) “Transitional Forms and the Evolution of Phyla“, Perspectives on Science and Christian Faith 53(1):42-51
Nielsen, Claus (2001) Animal Evolution: Interrelationships of the Living Phyla, Oxford University Press
Nielsen, Claus (2008) “Six major steps in animal evolution: are we derived sponge larvae?” Evolution & Development 10(2):241-257 (abstract)
Peterson, Kevin J. & Butterfield, Nicholas J. (2005) “Origin of the Eumetazoa: Testing ecological predictions of molecular clocks against the Proterozoic fossil record“, Proceedings of the National Academy of Science, 102(27):9547-9552
Porter, S. M. (2006) “The Proterozoic fossil record of heterotrophic protists“, In Xiao, S., and Kaufman, A.J. (eds.), Neoproterozoic Geobiology. Geobiology Series.
Ruiz-Trillo, Iñaki et al (1999) “Acoel Flatworms: Earliest Extant Bilaterian Metazoans, Not Members of Platyhelminthes”, Science 283(5409):1919-1923 (abstract)
Sedgwick, Adam (1909) A Student’s Text-Book of Zoology, Vol. III, Swan Sonnenschein and Company LTD, London/The Macmillan Company, New York.
Valentine, James W. (2004) On the Origin of Phyla, University of Chicago Press
Waggoner, Ben (2001) “Eukaryotes and Multicells: Origin” in Encyclopedia of Life Sciences (2001) Macmillan Publishers Ltd, Nature Publishing Group

Wow! This is a lovely summary of evolutionary signposts. I want to drop it entire into my blog, but I’ll content myself with linking to it. Way to go!
LikeLike
Pingback: Take that, creationists! « Science Notes
A positively terrific analysis of John Morris’ enduringly myopic grasp (or more accurately, lack of grasp) of biological evolution, and summary of the abundant diversity of quality evidence that drives and well-corroborates current evolution theory and thinking, Troy. I will be referring many others to your “tunicate” post, THANKS for the time, thought and work you put into it, you’ve produced a VERY nice piece of work!
LikeLike
Excellent work as usual Troy! I am thankful for your thorough rebutals to all the creationist crap that continues to linger.
LikeLike
Nice! A very thorough dismantling of creationist nonsense. Why do they go through all this trouble to bear false witness?
LikeLike
“Evolution Facts”; it’s really sad, but most Creationists, either wilfully or through simple lazy adherence to creed without any actual research, engage in the activity known as “Lying For Jesus”. Apparently someone in whatever organization they belong to (there are several, but it appears there was some coordination between them) decided that LFJ is perfectly acceptable behavior when dealing with “Satanistic, evil, baby-eating and possibly homosexual secular scientists”.
LikeLike
Dr. Morris chose a 44 year old popular level book from the Time-Life Nature Library series.
Hey! I remember that one! Heck, it may even still be somewhere in the house, since I know I still have others of the series. They were great when I was 10-15 years old … which no doubt explains why Dr. Morris chose it.
LikeLike
Well done! This would have been painful if I didn’t hate these lunatics so much. It was like watching a fistfight between a child and a grown man.
Don’t know whether you were aware, but this guy’s father Henry M. Morris wrote a wacked-out racist book called The Beginning of the World. He’s one of those guys who thinks the so-called “races” were “created” separately. Here’s a blog I did about it, and creationist racism in general. Wonder if that’s what John believes as well?
LikeLike
EXCELLENT job in deconstructing and exposing creationist mendacity and in educating us with your fine scientific research/detective work. You’ve done yeoman’s service here!
~David D.G.
LikeLike
It seems to me……….
If one was to beleive that god created all life on earth at relatively the same time then the quantity of species just after the creation should be equal to the number of years since the creation times the average rate of extinctions per year plus all present, living species.
The end result has got to be rediculesly unreal. The total number of species alive today has to be a slight fraction of the begining quantity.
Some one should plug some numbers into this simple creationist formula and see what happens. ……….and they say that the theory of evolution has gaps………..
LikeLike
Pingback: More scientific ignorance from Dr. John Morris « Playing Chess with Pigeons
Pingback: ‘O Oysters, come and walk with us! « Playing Chess with Pigeons
Hi
You are clearly very knowledgable in the field of biology.
How do we answer the scientifically accurate criticism that all creatures, single cell, multicell, vertibrates, invertibrates etc. can only reproduce their own species within the confines of their own gene pool. That mutations are always either useless or detremental, and natural selection (survival of the fittest) only serves to reduce variety in species.
Decades of experiments and observations seem to have well established these facts.
Please give, as far as possible a simple reply, as I am an engineer not a biologist, but am very interested in these things.
Kind Regards
Martin
LikeLike
Oh man, I don’t think so!
LikeLike
It is probably easier to teach a pidgeon chess at a grandmaster level than it is to teach a creationist think to about religion in a logical and sensible manner.
Q. How many creationists does it take to change a light bulb?
A. One million and eleven.
One to change the bulb when no one is looking.
Ten to make sure no one is looking.
And a million to claim the light bulb is eternal and can never burn out.
LikeLike
Pingback: Epic Horse Exhaust « Playing Chess with Pigeons
Pretty interesting post.
LikeLike
Pingback: There’s something fishy about that fish « Playing Chess with Pigeons
Pingback: Open mouth, insert hoof « Playing Chess with Pigeons
Great post! Thank you for taking the time to set this out so clearly.
LikeLike
Thank you and your welcome.
LikeLike
Pingback: Round 2 of: What “hope” do you have being an evolutionist? | Playing Chess with Pigeons